

«Grau, teurer Freund, ist alle Theorie
und grün des Lebens gold’ner Baum«
«Grey, dear friend, is all theory
and green life’s golden tree»
Johann Wolfgang Goethe,
Translation by David Marjanović
Humans are natural taxonomists. Whether a «lumper» or a «splitter» a person will invariably classify the things around them into logical groups and then refer to these groups to discuss a large range of objects rather than one, specifically. Scientists go through the same processes when classifying organisms, but whereas common (of folk) classifications spring from human perspectives, formal taxonomy is (or, at least, strives to be) an objective assessment of an organism’s actual relationships. Folk classifications are eminently serviceable to their own purposes, and often very complex, but the classification system used by modern biologists (cladistic taxonomy) is based upon different criteria. To construct a cladogram, a picture of «life’s golden tree», one must be able to separate true relationships between organisms from mere superficial similarities. Exactly how one goes about doing that requires some background and is the subject of this introductory chapter.
EVOLUTION
By the 18th and 19th centuries, zoologists were beginning to draw conclusions (or at least ask better questions) about life’s origins. As European scholars set out to systematically catalogue natural world, they began to notice patterns. Elephants in Africa bore a striking resemblance to those in India, but kangaroos in Australia looked like nothing else on Earth. Just as humans had bred different kinds of dogs, there was clearly some process was at work, making different breeds of everything else. For most of history, scholars, if they gave the matter any thought, followed Plato in assuming that modern breeds of organisms were corrupted descendants of perfect archetypes. Later biologists (most notably Jean de Monet, the Chevalier de Lamarck) theorized that an organism’s environment communicated changes that it could pass on to the next generation. We are all familiar with the example of the giraffes stretching their necks and the smith’s strong right arm being passed to his son, but these early evolutionary hypotheses do have some merit. The Platonic ideal of corrupted archytypes (if one strips away the moral connotations) are strikingly similar to Darwinian theory (which I will get to in a minute) and Lamarckian evolution, long considered completely wrong-headed, has now been revealed as a valid method of biological change, at least in some cases. Modern evolutionary theory, however, is most emphatically not Lamarckian or Platonic, but Darwinian.
Charles Darwin was (by a very short margin) the first person to publish a description of natural selection. Natural selection is nothing more than the automatic version of the selective breeding that humans have practiced for thousands of years, and is probably the most basic mechanism by which species change over time.
Very briefly, Darwinian evolution functions by killing every member of a population that does not conform to certain standards. That is, those that cannot survive do not. Whether by accident or design, genetic variation enters the system, and one organism ends up a little different from another. Some variations are beneficial, increasing the organism’s chances of reproduction, others are neutral, and some are detrimental, making the organism less likely to breed (by, for instance, killing it). Those organisms that do survive will hopefully breed, and pass on whatever trait kept them alive. Straightforward Darwinian evolution is very slow, with beneficial mutations building up over the eons and, little by little, creating a new species. Evolution by natural selection is simple, but the process is subject to much elaboration.
Sometimes evolution is neither simple nor slow. If, for instance, two populations of the same species are isolated so individuals of the two populations can no longer interbreed, they will begin to build up distinct mutations. These mutations may not be particularly beneficial—two spots on the wings rather than three, or blue eyes instead of green eyes—but enough of them will make the two populations so different from each other that they become different species. Even if the barrier between the two populations were removed, they would be unable to interbreed.
Sexual selection adds another level of complication. Females will often favor males who sport obvious disatvantages—conspicuous coloration, large, biologically expensive horns, or prodigious scars—the reasoning being that any male with such tremendous handicaps must be very fit to be able to survive at all. Masculine ornamentation (females are too valuable to waste their resources on such frippery) thus becomes a barometer for their general health. As females select males with the best ornamentation, many species find themselves with the fantastically decorated men-folk; the gaudy plumage of a carpo-of-paradise, the great antlers of a hornmeister, or the scarred death-mask of a ravana (the product of many victories) are all the products of thousands of generations of sexual selection.
Another interesting twist on classical Darwinian theory is that genetic variation need not be expressed as a change in the organism’s form. Most of our DNA is not used at all, and functions as a source of backup variety to be used when the right circumstances present themselves. If a number of organisms find themselves in an environment with few competitors, these organisms go through an evolutionary radiation and diversify with dazzling speed. The cichlids of Africa’s lake Victoria, for instance, could not have been living there more than 750,000 years (the lake, itself is at most that old), but in this eyeblink of time, they have radiated into an array of species to cover almost every lifestyle a fish could have. Genetic tests conducted by Axel Meyer et al. indicate these different cichlids species actually contain less genetic variety than the single species of Homo sapiens. The difference between us and the fish is that while most humans’ genetic variation is latent, the cichlids have expressed their genetic diversity physically.
There are only so many different environments on Earth, and as species adapt to their surroundings, the same adaptations are re-invented again and again. Convergent evolution occurs when two species of wholly different lineage share an outward appearance molded by their environment. Spec abounds with examples of convergence (as does Home-Earth), some of which include the twitiavians mistriders versus the neognath swoops or the North American nodopotami [1] versus the Eurasian potamoceratopians. In both these cases, the outward form of two groups of organisms (or taxa) are similar even though they are not related. The only way to sort through convergences is to examine those parts of the organisms’ anatomies that not subject to the selective pressures of their environments or cannot be changed if they are. A mistrider, for instance, though it may look like a swift, possesses the bizarre palate configuration of a twitiavian. The fact that the mistriders look nothing at all like other twitiavians is due to divergent evolution , where a different style of life has lead to a different body type from its ancestors.
Another form evolutionary change, neither divergent nor exactly convergent, is parallel evolution, observed where Spec-world taxa have evolved in parallel with their counterparts Arel counterparts. Most high-level taxa are parallel (that is, phylum Chordata exists both on the Spec-world and Home-Earth), but there are a surprising number of genus and even species-level examples of parallelism.
Evolution as a concept may be simple, but the tremendous diversity of life both on Spec and Home-Earth is testament to the complexity the underlies natural-selections elegance. Species drift apart, radiate, diverge, converge, and generally make the process of tracing back their lineages that much harder.
CLASSIFICATION
Life has been evolving for almost as long as the Earth has been solid (some three billion years) and, evolution being a build-up of many small changes, some organisms are quite different from others. Other organisms, however, are more similar, and some are quite similar. Logically, those organisms most like each other are the most closely related, having had less time to develop distinction. This argument, in a nutshell, is what cladistic taxonomy is all about.
Cladistics (from Greek klados, twig) classifies organisms by arranging them into branching trees, where the most closely related species are physically closest together. Cladistics (or at least, the classification of organisms) traces its own ancestry back to the system devised by Carl von Linné (Carolus Linnaeus) in the 1700s. Linnaean classification was mostly concerned with naming organisms and placing them in hierarchies based upon their similarities. The mosarks that hunt the upper levels of the ocean, for instance constitute a species, a community of animals that can interbreed to produce offspring with can then breed, themselves. This species belongs to a larger (and somewhat arbitrary) grouping, a genus of other animals which are very much like the common mosark but cannot breed with this species. These two names, genus and species, form the Linnaean name of an organism. Linnaean names are usually Greek or Latin (though not always) and are written in italics, with the first letter of the generic name capitalized. Thus, the Linnaean name of the common mosark is Pristrix monstrum. Species borders are sometimes fuzzy, but in general, every P. monstrum is genetically compatible with every other P. monstrum, but not with any other organism, though those of genus Pristrix (the blue-striped mosark or Pristrix coloris, for instance) may look similar. Linnaeus created a system whereby every organism on Earth was designated by a pair of names, generic and specific. No two species have the same Linnaean name; there are elaborate rules set up to ensure that no such confusion may occur.
Linnaeus invented more than the idea of the species however, he also set up a complex hierarchy into which species could be placed. The common mosark belongs to the species Pristrix monstrum, the genus Pristrix, the family Mosasauria (or Mosasauridae, if one is being technical), the order Squamata, class Reptilia, phylum Chordata, kingdom Animalia, and domain Eucarya. Each of these groupings — taxa (singular taxon) — are based upon one or more specific characteristics (traits) that all species within the taxa share with each other and not with any other organisms. For instance, Linnaeus grouped all creatures with feathers into class Aves and all creatures with hair and nipples into class Mammalia. If the Swedish biologist had lived in Spec, where feathered dinosaurs and hairy, nipple-less multituberculates are common, he would probably have drawn his classifications differently, but that is beside the point.
Logical as it is, there are problems, with the Linnaen system. Most fundamental is the fact that by Carl von Linné was born before Charles Darwin published The Origin of Species and introduced the world to the concept of evolution through natural selection. Linnaeus used Plato’s method to explain the resemblance of some organisms to others, and said that modern creatures were the corrupt descendants of a set of perfect archetypes. Darwin, took this idea even further in saying that all life on Earth is descended from a single species. Now, the purely arbitrary hierarchies of Linnaeus could be viewed as actual family trees, describing evolutionary changes through time. How, though, can one describe the infinitely subtle variations of groups of species with just the few arbitrary hierarchies set up by Linnaeus? Of course, some changes to the old system had to be made.
Cladistics retains much of the old Linnaen terminology. Species are still defined as interbreeding populations and given binomial titles. In this book, we even use such designations as «family» and «order» to describe groupings of species. How, though, can one group together genera within an family, or families within an order? Since Linnaeus, biologists have created a number of intermediate groupings like «subclasses» and «infraorders» for this purpose. Such a process could potentially extend indefinably, with biologists naming «subsubclasses», «supersubphyla», or «subsuperinfraorders» to classify species with greater and greater precision. What is more, if one believes taxonomy should be based upon evolutionary descent, then classes Mammalia and Aves should both be inserted into class Reptilia, since both mammals and birds evolved from reptiles. Furthermore, with convergence making unrelated organisms look the same, and divergence making related organisms look different, who can tell how anything is related to anything else?
Cladists wrestled with these problems (among others) through most of the 20th century, and emerged with a system, that while not perfect, provides some solutions.
The fundamental idea of cladistics is that features shared by organisms because they share common ancestors (plesiomorphies and apomorphies), rather than features shared by organisms because of convergence or coincidence (homoplasies), should be used to construct family trees. For instance, a mosark and a tiger shark share several features (called «characters» in cladospeak). Both creatures are bullet-shaped, with a set of fins are sharp teeth. However, these characters are all homoplasies («the same molds») they arose independently in both sharks and mosarks as adaptations to the lifestyle of a hunter of the open ocean. The shark is not closely related to the shark, it turns out. The tiger shark is a cartilaginous fish, a specifically a lamniform chondrichtyan, while the mosark is a lizard, a mosaurian squamatan. Other features shared by these two organisms are not homoplasious, however. Both sharks and mosarks share eyes with pupils, hinged jaws, and a series of vertebrae funning down the back to form the spine or backbone. All these characters (and many more) are shared by the shark and the mosark because both organisms evolved from a common ancestor; the characters are plesiomorphies («near-forms»). If one then brings another organism into this group, say the boa, a new series of shared characters will spring up. The boa, as a vertebrate, will share many plesiomorphies with both the shark and the mosark. However, the observer will note some further similarities shared between the boa and the mosark, but not by the shark. Both the snake and mosasaur share a bony (not cartilaginous) skeleton, five digits per limb [2] (although the mosark’s fingers and toes are fused into paddles), and a unique skull with flexible joints that lend a great deal of mobility and strength to the jaw. These characters are shared/advanced, not shared/primative, and are called apomorphies («far-forms»). From this mix of homoplasies, plesiormorphies, and apomorphies the observer would conclude (and rightly so) that the boa and the mosark families split away from each other later than they did from the shark lineage.
Cladospeak is all well and good, but it is difficult to comprehend as words only. Biologists show the relationships between creatures as family trees, called cladograms («branch-pictures»). Graphically, these trees are based upon «sister taxa»—pairs of groupings (species, genera, etc.) that share some distinctive characters. The difference between this cladistic classification and a Linnaean hierarchy is that a cladogram is fractal, it looks the same at all levels, with sister species, sister genera, sister families, and so on, all the way to sister domains, the most basic classification of life.
To use the example of the tiger shark, mosark, and boa, a cladogram would show the mosark and boa as sister taxa, since they share a number of apomorphies and plesiomorphies, the resulting cladogram would look like this:
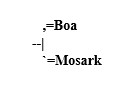
If one then adds the tiger shark to this picture, one sees that the shark shares plesiormophies with the boa and the mosark, but does not share any apomorphies with either. Thus, the shark is a sister taxon to the group boa+mosark:

Referring to taxa as boa+mosark, while technically correct, can get cumbersome, so taxonomists arbitrarily name various groups of taxa (these groups can be called taxa, as well, since cladistics is fractal) as clades. Many of these clades are inherited from Linnean taxonomy, and so tend to form a hierarchy of phyla, orders, and so on. This hierarchy (aside from species) does not reflect any fundamental reality of an organism’s relationship to other organisms, but it is useful as a codofier:

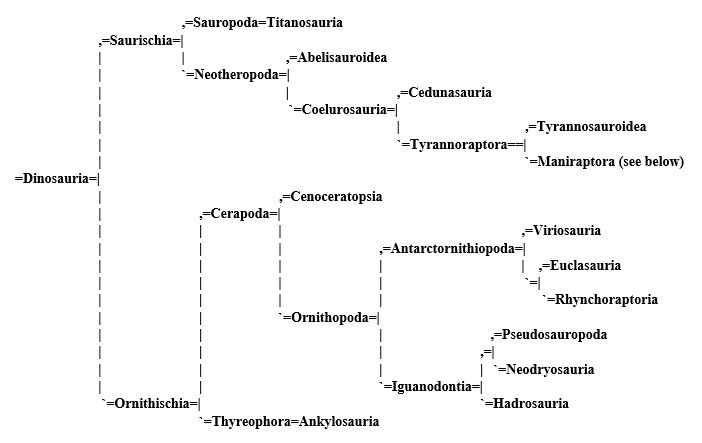
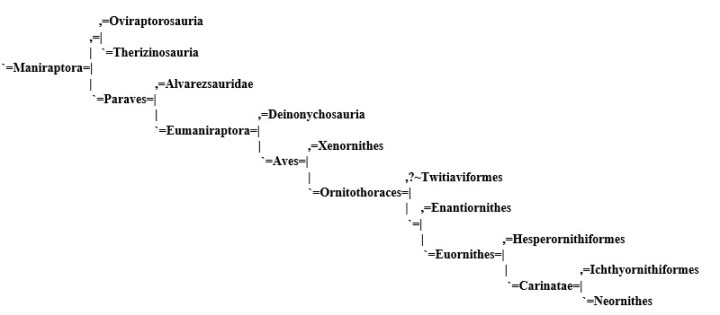
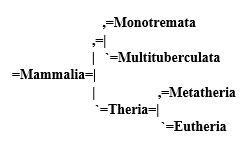
The exact placement of these groups (and their names) is subject to much argument and is dictated by a series of byzantine rules dating back more than a century, but the basic rules of cladistics as we put them, stand. All of the cladograms of this project were constructed according this same principals, and at a glance will show the relationships between the organisms we describe.
Text by Daniel Bensen.
THE CLADOGRAMS
Dinosaurs [3]:
Mammals:
Works referenced:
1. The Variety of Life by Colin Tudge. Published in 2000 by Oxford University Press.
2. The Tree of Life
3. The Vertebrate Phylogeny Pages
4. The Origin and Evolution of Birds (second edition) by Alan Feduccia. Published in 1999 by Yale University Press.
5. Manual of Ornithology by Noble S. Proctor & Patrick J, Lynch. Published in 1993 by Yale University Press.
Thanks to David Marjanović for his help with cladotalk.
Editor’s note:
[1] Removed in the later versions of Spec.
[2] Obviously snakes do not have limbs, nor digits. This trait, however, was acquired after they splitted from mosasaurs.
[3] Here Twitiaviformes are shown outside Enentiornithes, which reflects the earlier versions of Spec.


